Bazálne gangliá telencefalu. Bazálne gangliá
Pohyb a myslenie sú vlastnosti, ktoré umožňujú človeku naplno žiť a rozvíjať sa.
Aj malé poruchy mozgových štruktúr môžu viesť k významným zmenám alebo úplnej strate týchto schopností.
Za tieto nevyhnutné životné procesy sú zodpovedné skupiny nervových buniek v mozgu nazývané bazálne gangliá.
Čo potrebujete vedieť o bazálnych gangliách
Veľké hemisféry ľudského mozgu na vonkajšej strane sú kôra tvorená sivou hmotou a zvnútra subkortexom bielej hmoty. Bazálne gangliá (ganglia, uzliny), ktoré sa tiež nazývajú centrálne alebo subkortikálne, sú koncentrácie šedej hmoty v bielej hmote subkortexu.
Bazálne gangliá sa nachádzajú v spodnej časti mozgu, čo vysvetľuje ich názov, mimo talamu (optický talamus). Ide o párové útvary, ktoré sú symetricky zastúpené v oboch hemisférach mozgu. Pomocou nervových procesov interagujú bilaterálne s rôznymi oblasťami centrálnej nervový systém.
Hlavnou úlohou subkortikálnych uzlov je organizovať motorickú funkciu a rôzne aspekty vyššia nervová aktivita. Patológie, ktoré vznikajú v ich štruktúre, ovplyvňujú fungovanie iných častí centrálneho nervového systému, spôsobujú problémy s rečou, koordináciou pohybov, pamäťou a reflexmi.
Vlastnosti štruktúry bazálnych ganglií
Bazálne gangliá sa nachádzajú v predných a čiastočne temporálnych lalokoch telencefalu. Ide o zhluky telies neurónov, ktoré tvoria skupiny šedej hmoty. Biela hmota, ktorá ich obklopuje, je reprezentovaná procesmi nervových buniek a vytvára vrstvy, ktoré oddeľujú jednotlivé bazálne gangliá a ďalšie štrukturálne a funkčné prvky mozgu.
Bazálne uzliny zahŕňajú:
- striatum;
- plot;
- amygdala.
Na anatomických rezoch sa striatum javí ako striedajúce sa vrstvy šedej a bielej hmoty. Pozostáva z kaudátových a lentikulárnych jadier. Prvý sa nachádza v prednej časti vizuálneho talamu. Keď sa caudate nucleus stáva tenším, stáva sa z neho amygdala. Lentikulárne jadro je umiestnené laterálne od talamu opticus a nucleus caudatus. Je s nimi spojený tenkými mostíkmi neurónov.
Plot je úzky pás neurónov. Nachádza sa medzi lentikulárnym jadrom a ostrovčekovou kôrou. Od týchto štruktúr je oddelená tenkými vrstvami bielej hmoty. Amygdala má tvar amygdaly a nachádza sa v spánkových lalokoch telencephalonu. Skladá sa z niekoľkých nezávislých prvkov.
Táto klasifikácia je založená na štrukturálnych znakoch a umiestnení ganglií na anatomickej časti mozgu. Existuje aj funkčná klasifikácia, podľa ktorej vedci zaraďujú medzi bazálne gangliá len striatum a niektoré gangliá diencefala a mezencefala. Tieto štruktúry spolu poskytujú motorické funkcie osoba a jednotlivé aspekty správania zodpovedné za motiváciu.
Anatómia a fyziológia bazálnych ganglií
Hoci všetky bazálne gangliá sú zbierkami šedej hmoty, majú svoj vlastný komplex štrukturálne vlastnosti. Aby sme pochopili, akú úlohu zohráva toto alebo toto bazálne centrum vo fungovaní tela, je potrebné bližšie sa pozrieť na jeho štruktúru a umiestnenie.
Caudate jadro
Tento subkortikálny uzol sa nachádza v predných lalokoch mozgových hemisfér. Je rozdelená do niekoľkých sekcií: zahustená veľká hlava, zužujúce sa telo a tenký dlhý chvost. Caudatus nucleus je vysoko predĺžený a zakrivený. Ganglion pozostáva väčšinou z mikroneurónov (do 20 mikrónov) s krátkymi tenkými výbežkami. Asi 5 % celkovej bunkovej hmoty subkortikálneho ganglia pozostáva z väčších nervových buniek (do 50 mikrónov) s vysoko rozvetvenými dendritmi.
Tento ganglion interaguje s oblasťami kôry, talamu a uzlov diencefala a stredného mozgu. On hrá rolu styčný medzi týmito mozgovými štruktúrami, pričom neustále prenáša nervové impulzy z mozgovej kôry do jej ostatných častí a späť. Je multifunkčný, no jeho úloha je významná najmä pri udržiavaní činnosti nervovej sústavy, ktorá reguluje činnosť vnútorných orgánov.
Lentikulárne jadro
Tento bazálny uzol má tvar semien šošovice. Nachádza sa tiež v predných oblastiach mozgových hemisfér. Pri rezaní mozgu vo frontálnej rovine túto štruktúru je trojuholník, ktorého vrchol smeruje dovnútra. Biela hmota rozdeľuje tento ganglion na putamen a dve vrstvy globus pallidus. Škrupina je tmavá a umiestnená zvonka vo vzťahu k svetlým vrstvám globus pallidus. Neurónové zloženie putamenu je podobné nucleus caudatus, ale globus pallidus je reprezentovaný hlavne veľkými bunkami s malými inklúziami mikroneurónov.
Evolučne je globus pallidus uznávaný ako najstarší útvar spomedzi ostatných bazálnych ganglií. Putamen, globus pallidus a caudate nucleus tvoria striopallidálny systém, ktorý je súčasťou extrapyramídového systému. Hlavnou funkciou tohto systému je regulácia vôľových pohybov. Anatomicky je spojená s mnohými kortikálnymi poľami mozgových hemisfér.
Plot
Mierne zakrivená, stenčená doska šedej hmoty, ktorá oddeľuje putamen a insula telencephalon, sa nazýva plot. Biela hmota okolo tvorí dve kapsuly: vonkajšiu a „vonkajšiu“. Tieto kapsuly oddeľujú plot od susedných štruktúr šedej hmoty. Plot prilieha k vnútornej vrstve neokortexu.
Hrúbka plotu sa pohybuje od zlomkov milimetra až po niekoľko milimetrov. Po celej dĺžke sa skladá z neurónov rôznych tvarov. Plot je spojený prostredníctvom nervových dráh s centrami mozgovej kôry, hipokampu, amygdaly a čiastočne striatálnych telies. Niektorí vedci považujú plot za pokračovanie mozgovej kôry alebo ho zahŕňajú ako súčasť limbického systému.
Amygdala
Tento ganglion je skupina buniek šedej hmoty sústredených pod škrupinou. Amygdala pozostáva z niekoľkých útvarov: jadrá kôry, stredné a centrálne jadrá, bazolaterálny komplex a intersticiálne bunky. Nervovým prenosom je spojený s hypotalamom, talamom, zmyslovými orgánmi, jadrami hlavových nervov, čuchovým centrom a mnohými ďalšími útvarmi. Niekedy je amygdala klasifikovaná ako súčasť limbického systému, ktorý je zodpovedný za činnosť vnútorných orgánov, emócie, vôňu, spánok a bdenie, učenie atď.
Význam subkortikálnych uzlín pre telo
Funkcie bazálnych ganglií sú určené ich interakciou s inými oblasťami centrálneho nervového systému. Tvoria nervové slučky spájajúce talamus a najdôležitejšie oblasti mozgovej kôry: motorickú, somatosenzorickú a frontálnu. Okrem toho sú subkortikálne uzliny spojené navzájom a s niektorými oblasťami mozgového kmeňa.
Caudate nucleus a putamen vykonávajú tieto funkcie:
- kontrola smeru, sily a amplitúdy pohybov;
- analytická činnosť, učenie, myslenie, pamäť, komunikácia;
- kontrola pohybov očí, úst a tváre;
- udržiavanie fungovania vnútorných orgánov;
- podmienená reflexná aktivita;
- vnímanie zmyslových signálov;
- kontrola svalového tonusu.
Špecifické funkcie škrupiny zahŕňajú dýchacie pohyby, produkciu slín a ďalšie aspekty kŕmenia, zabezpečujúce trofizmus kože a vnútorných orgánov.
Funkcie globus pallidus:
- vývoj orientačnej reakcie;
- kontrola pohybov rúk a nôh;
- stravovacie správanie;
- výrazy tváre;
- prejav emócií;
- poskytovanie pomocných pohybov a koordinačných schopností.
Funkcie plotu a amygdaly zahŕňajú:
- reč;
- stravovacie správanie;
- emocionálna a dlhodobá pamäť;
- vývoj behaviorálnych reakcií (strach, agresia, úzkosť atď.);
- zabezpečenie sociálnej integrácie.
Veľkosť a stav jednotlivých bazálnych ganglií teda ovplyvňuje emocionálne správanie, vôľové a mimovoľné pohyby človeka, ako aj vyššiu nervovú aktivitu.
Choroby bazálnych ganglií a ich symptómy
Narušenie normálneho fungovania bazálnych ganglií môže byť spôsobené infekciou, úrazom, genetickou predispozíciou, vrodenými anomáliami alebo metabolickým zlyhaním.
Symptómy patológie sa niekedy objavujú postupne, bez povšimnutia pacienta.
Mali by ste venovať pozornosť nasledujúcim príznakom:
- celkové zhoršenie zdravia, slabosť;
- zhoršený svalový tonus, obmedzený pohyb;
- výskyt dobrovoľných pohybov;
- chvenie;
- zhoršená koordinácia pohybov;
- výskyt neobvyklých pozícií pre pacienta;
- ochudobnenie výrazov tváre;
- zhoršenie pamäti, zakalenie vedomia.
Patológie bazálnych ganglií sa môžu prejaviť v mnohých ochoreniach:
- Funkčný nedostatok. Prevažne dedičné ochorenie, prejavujúce sa v detstva. Hlavné príznaky: neovládateľnosť, nepozornosť, enuréza do 10–12 rokov, nevhodné správanie, nejasné pohyby, zvláštne držanie tela.
- Cyst. Bez včasného lekárskeho zásahu vedú zhubné nádory k invalidite a smrti.
- Kortikálna paralýza. Hlavné príznaky: mimovoľné grimasy, zhoršená mimika, kŕče, chaotické pomalé pohyby.
- Parkinsonova choroba. Hlavné príznaky: tremor končatín a tela, znížená motorická aktivita.
- Huntingtonova choroba. Genetická patológia, ktorá postupuje postupne. Hlavné príznaky: spontánne nekontrolované pohyby, strata koordinácie, znížená mentálne schopnosti, depresie.
- . Hlavné príznaky: spomalenie a ochudobnenie reči, apatia, nevhodné správanie, zhoršenie pamäti, pozornosti a myslenia.
Niektoré funkcie bazálnych ganglií a znaky ich interakcie s inými štruktúrami mozgu ešte neboli stanovené. Neurológovia pokračujú v štúdiu týchto subkortikálnych centier, pretože ich úloha pri udržiavaní normálneho fungovania ľudského tela je nespochybniteľná.
V článku budeme hovoriť o bazálnych gangliách. Čo to je a akú úlohu zohráva táto štruktúra v ľudskom zdraví? Všetky otázky budú podrobne diskutované v článku, po ktorom pochopíte dôležitosť absolútne každého „detailu“ vo vašom tele a hlave.
o čom to hovoríme?
Všetci veľmi dobre vieme, že ľudský mozog je veľmi zložitá jedinečná štruktúra, v ktorej sú absolútne všetky prvky neoddeliteľne a pevne spojené prostredníctvom miliónov nervových spojení. V mozgu je šedá a prvá je bežná akumulácia mnohých nervových buniek a druhá je zodpovedná za rýchlosť prenosu impulzov medzi neurónmi. Okrem kôry samozrejme existujú aj iné štruktúry. Sú to jadrá alebo bazálne gangliá, ktoré pozostávajú zo šedej hmoty a nachádzajú sa v bielej hmote. V mnohých ohľadoch sú zodpovedné za normálne fungovanie nervového systému.
Bazálne gangliá: fyziológia
Tieto jadrá sa nachádzajú v blízkosti mozgových hemisfér. Majú veľa dlhých procesov nazývaných axóny. Vďaka nim sa informácie, teda nervové impulzy, prenášajú do rôznych štruktúr mozgu.
Štruktúra
Štruktúra bazálnych ganglií je rôznorodá. V podstate sa podľa tejto klasifikácie delia na tie, ktoré patria do extrapyramídového a limbického systému. Oba tieto systémy majú obrovský vplyv na fungovanie mozgu a sú s ním v úzkej interakcii. Ovplyvňujú talamus, parietálny a čelný lalok. Extrapyramídová sieť pozostáva z bazálnych ganglií. Kompletne prekrvuje podkôrové časti mozgu a má zásadný vplyv na fungovanie všetkých funkcií ľudského tela. Tieto skromné útvary zostávajú veľmi často podceňované, a napriek tomu ich práca ešte nie je úplne preštudovaná.

Funkcie
Funkcií bazálnych ganglií nie je veľa, ale sú významné. Ako už vieme, sú silne prepojené so všetkými ostatnými mozgovými štruktúrami. V skutočnosti z pochopenia tohto tvrdenia vyplývajú hlavné:
- Kontrola nad realizáciou integračných procesov pri vyššej nervovej činnosti.
- Vplyv na fungovanie autonómneho nervového systému.
- Regulácia motorických procesov človeka.
Do čoho sa zapájajú?
Existuje množstvo procesov, na ktorých sa priamo podieľajú jadrá. Bazálne gangliá, ktorých štruktúra, vývoj a funkcie uvažujeme, sa podieľajú na nasledujúcich akciách:
- ovplyvniť zručnosť osoby pri používaní nožníc;
- presnosť zatĺkania klincov;
- reakčná rýchlosť, driblovanie s loptou, presnosť zásahu do koša a obratnosť zásahu loptou pri hraní basketbalu, futbalu, volejbalu;
- ovládanie hlasu pri speve;
- koordinácia akcií pri kopaní zeme.
Tieto jadrá ovplyvňujú aj zložité motorické procesy, ako je jemná motorika. To je vyjadrené v spôsobe, akým sa ruka pohybuje pri písaní alebo kreslení. Ak dôjde k narušeniu fungovania týchto mozgových štruktúr, potom bude rukopis nečitateľný, hrubý a „neistý“. Inými slovami, bude sa zdať, že osoba len nedávno vzala do ruky pero.

Nový výskum ukázal, že bazálne gangliá môžu tiež ovplyvniť typ pohybu:
- kontrolovateľné alebo náhle;
- mnohokrát opakované alebo nové, úplne neznáme;
- jednoduché jednoslabičné alebo postupné a dokonca simultánne.
Mnohí výskumníci, nie bezdôvodne, veria, že funkcie bazálnych ganglií spočívajú v tom, že človek môže konať automaticky. To naznačuje, že veľa akcií, ktoré človek vykonáva na cestách, bez toho, aby im venoval pozornosť osobitnú pozornosť, sú možné práve vďaka jadrám. Fyziológia bazálnych ganglií je taká, že riadia a regulujú ľudské automatické aktivity bez odoberania zdrojov z centrálneho nervového systému. To znamená, že musíme pochopiť, že práve tieto štruktúry do veľkej miery kontrolujú, ako človek koná v strese alebo v nepochopiteľnej nebezpečnej situácii.
IN bežný život bazálne gangliá jednoducho prenášajú impulzy, ktoré prichádzajú z predných lalokov, do iných štruktúr mozgu. Cieľom je cielene vykonávať známe činnosti bez stresu centrálneho nervového systému. V nebezpečných situáciách sa však gangliá „prepnú“ a umožnia človeku automaticky urobiť najoptimálnejšie rozhodnutie.

Patológie
Lézie bazálnych ganglií môžu byť veľmi odlišné. Pozrime sa na niektoré z nich. Ide o degeneratívne lézie ľudského mozgu (napríklad Parkinsonova choroba alebo Huntingtonova chorea). Môže ísť o dedičné genetické ochorenia, ktoré sú spojené s metabolickými poruchami. Patológie charakterizované poruchami enzýmových systémov. Choroby štítnej žľazy sa môžu vyskytnúť aj v dôsledku porúch vo fungovaní jadier. Možné patológie vyplývajúce z otravy mangánom. Mozgové nádory môžu ovplyvniť fungovanie bazálnych ganglií a to je možno tá najnepríjemnejšia situácia.

Formy patológií
Vedci bežne identifikujú dve hlavné formy patológie, ktoré sa môžu vyskytnúť u ľudí:
- Funkčné problémy. Toto sa často vyskytuje u detí. Príčinou je vo väčšine prípadov genetika. Môže sa vyskytnúť u dospelých po mŕtvici, ťažkej traume alebo krvácaní. Mimochodom, v starobe sú to poruchy ľudského extrapyramídového systému, ktoré spôsobujú Parkinsonovu chorobu.
- Nádory a cysty. Táto patológia je veľmi nebezpečná a vyžaduje okamžitú lekársku intervenciu. Charakteristickým príznakom je prítomnosť závažných a dlhotrvajúcich neurologických ochorení.
Za zmienku tiež stojí, že bazálne gangliá mozgu môžu ovplyvniť flexibilitu ľudského správania. To znamená, že človek sa v ňom začne strácať rôzne situácie nedokážu rýchlo reagovať, prispôsobiť sa ťažkostiam alebo jednoducho konať podľa svojho obvyklého algoritmu. Je tiež ťažké pochopiť, ako logicky konať v situácii, ktorá je pre normálneho človeka jednoduchá.
Poškodenie bazálnych ganglií je nebezpečné, pretože človek sa stáva prakticky nepoučiteľným. Je to logické, pretože učenie je podobné automatizovanej úlohe a ako vieme, tieto jadrá sú za takéto úlohy zodpovedné. Je však liečiteľná, aj keď veľmi pomaly. V tomto prípade budú výsledky zanedbateľné. Na tomto pozadí človek prestáva ovládať svoju koordináciu pohybov. Zvonku sa zdá, že sa pohybuje prudko a prudko, akoby sebou trhal. V tomto prípade môže skutočne dôjsť k chveniu končatín alebo k nejakým mimovoľným úkonom, ktoré pacient nemá pod kontrolou.

Oprava
Liečba poruchy úplne závisí od toho, čo ju spôsobilo. Liečbu vykonáva neurológ. Veľmi často sa problém dá vyriešiť len pomocou neustálej medikácie. Tieto systémy nie sú schopné obnovy samy o sebe, ale tradičné metódy sú mimoriadne zriedkavé. Hlavná vec, ktorá sa od človeka vyžaduje, je konzultovať s lekárom včas, pretože len to zlepší situáciu a dokonca sa vyhne veľmi nepríjemným príznakom. Lekár stanoví diagnózu pozorovaním pacienta. Tiež používané moderné metódy diagnostika ako MRI a CT mozgu.
Zhrnutím článku by som chcel povedať, že na bežnú prevádzku ľudské telo, a najmä mozgu, je veľmi dôležité správne fungovanie všetkých jeho štruktúr a aj tých, ktoré sa na prvý pohľad môžu zdať úplne bezvýznamné.
Shell
Bledá lopta
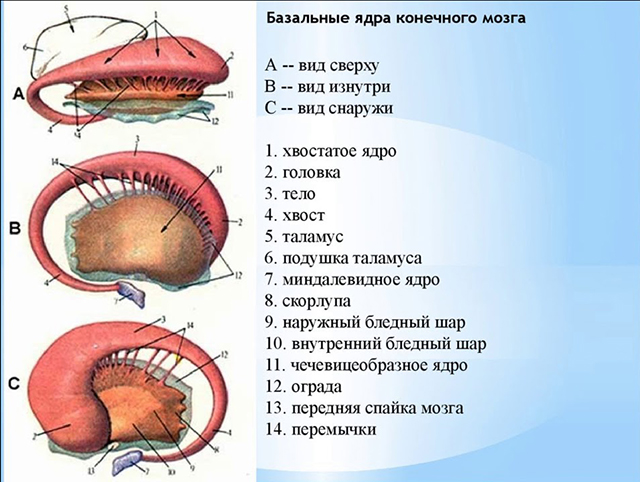
V hrúbke bielej hmoty každej hemisféry veľký mozog sú akumulácie šedej hmoty tvoriace samostatne ležiace jadrá (obr. 7). Tieto jadrá ležia bližšie k základni mozgu a nazývajú sa bazálne (subkortikálne, centrálne). Patria sem: 1) pruhované telo, ktoré u nižších stavovcov tvorí prevládajúcu hmotu hemisfér; 2) plot; 3) amygdala.
Uvažujme o štruktúre striata (corpus striatum), ktoré v častiach mozgu vyzerá ako striedajúce sa pruhy šedej a bielej hmoty. Najviac v strede a vpredu je: a) caudate nucleus, umiestnený laterálne a nad talamom, pričom je od neho oddelený kolenom vnútornej kapsuly. Jadro má hlavu umiestnenú v prednom laloku, vyčnievajúcu do predného rohu laterálnej komory a priliehajúcu k prednej perforovanej substancii. Telo caudatus nucleus leží pod parietálnym lalokom a ohraničuje centrálnu časť laterálnej komory na laterálnej strane. Chvost jadra sa podieľa na tvorbe strechy dolného rohu laterálnej komory a dosahuje amygdalu, ktorá leží v anteromediálnych častiach spánkového laloku (za prednou perforovanou látkou); b) lentikulárny jadro je umiestnené laterálne od nucleus caudate. Vrstva bielej hmoty - vnútorná kapsula– oddeľuje lentikulárne jadro od jadra caudatus a od talamu.
 Spodný povrch prednej časti lentiformného jadra susedí s prednou perforovanou substanciou a je spojený s nucleus caudate. Stredná časť lentikulárneho jadra v horizontálnom reze mozgu sa zužuje a je sklonená smerom ku kolenu vnútornej kapsuly, ktorá sa nachádza na hranici talamu a hlavy nucleus caudatus. Konvexný laterálny povrch lentikulárneho jadra smeruje k základni ostrovného laloku mozgovej hemisféry.
Spodný povrch prednej časti lentiformného jadra susedí s prednou perforovanou substanciou a je spojený s nucleus caudate. Stredná časť lentikulárneho jadra v horizontálnom reze mozgu sa zužuje a je sklonená smerom ku kolenu vnútornej kapsuly, ktorá sa nachádza na hranici talamu a hlavy nucleus caudatus. Konvexný laterálny povrch lentikulárneho jadra smeruje k základni ostrovného laloku mozgovej hemisféry.
Obr.7. Predná časť mozgu na úrovni mastoidných teliesok.
1 – cievnatka plexus laterálnej komory (centrálna časť), 2 – talamus, 3 – vnútorná kapsula, 4 – insular cortex, 5 – plot, 6 – amygdala, 7 – zraková dráha, 8 – mastoidné telo, 9 – globus pallidus, 10 – putamen, 11 – fornix, 12 – nucleus caudate, 13 – corpus callosum.
Lentikulárne jadro má na frontálnom úseku mozgu tiež tvar trojuholníka, ktorého vrchol smeruje k mediálnej strane a báza k laterálnej strane (obr. 7). Dve paralelné vertikálne vrstvy bielej hmoty rozdeľujú lentikulárne jadro na tri časti. Tmavší leží najviac laterálne škrupina, mediálnejšie je " bledá guľa“, pozostávajúce z dvoch dosiek: mediálnej a laterálnej. K fylogeneticky novším útvarom patrí caudate nucleus a putamen, k starším globus pallidus. Jadrá striata tvoria striopallidálny systém, ktorý zase patrí do extrapyramídového systému zapojeného do riadenia pohybov a regulácie svalového tonusu (obr.).
Obr.8. Horizontálna časť mozgu. Bazálne gangliá.
 1–mozgová kôra (plášť), 2–rod corpus callosum, 3–predný roh laterálnej komory, 4–vnútorná kapsula, 5–vonkajšia kapsula, 6–plot, 7–vonkajšia kapsula, 8–putamen, 9– globus pallidus , 10–III komora, 11–zadný roh laterálnej komory, 12–tuberkula zrakového nervu, 13–kortikálna substancia (kôra) insuly, 14–hlav.
1–mozgová kôra (plášť), 2–rod corpus callosum, 3–predný roh laterálnej komory, 4–vnútorná kapsula, 5–vonkajšia kapsula, 6–plot, 7–vonkajšia kapsula, 8–putamen, 9– globus pallidus , 10–III komora, 11–zadný roh laterálnej komory, 12–tuberkula zrakového nervu, 13–kortikálna substancia (kôra) insuly, 14–hlav.
Slim vertikálne umiestnený plot, ležiaci v bielej hmote hemisféry na strane obalu, je oddelený od obalu vonkajším puzdrom a od ostrovčekovej kôry najvzdialenejším puzdrom.
Caudate nucleus a putamen dostávajú zostupné spojenia primárne z extrapyramídového kortexu cez subkalosálny fasciculus. Iné polia mozgovej kôry tiež posielajú veľké množstvo axóny do nucleus caudate a putamen.
Hlavná časť axónov nucleus caudate a putamen smeruje do globus pallidus, odtiaľ do talamu a až z neho do zmyslových polí. V dôsledku toho medzi týmito formáciami existuje začarovaný kruh spojení. Caudatus nucleus a putamen majú tiež funkčné spojenie so štruktúrami ležiacimi mimo tohto kruhu: so substantia nigra, červeným jadrom, Lewisovým telom (subtalamické jadro), vestibulárnymi jadrami, mozočkom, gama bunkami miecha.
Množstvo a povaha spojení medzi caudate nucleus a putamen naznačuje ich účasť na integračných procesoch, organizácii a regulácii pohybov a regulácii práce vegetatívnych orgánov.
Mediálne jadrá talamu majú priame spojenie s nucleus caudatus, o čom svedčí reakcia jeho neurónov, ku ktorej dochádza 2-4 ms po stimulácii talamu. Reakcia neurónov v nucleus caudatus je spôsobená podráždením kože, svetelnými a zvukovými podnetmi.
Pri nedostatku dopamínu v nucleus caudatus (napríklad pri dysfunkcii substantia nigra) dochádza k dezinhibícii globus pallidus, aktivácii systémov chrbtice a drieku, čo vedie k motorickým poruchám v podobe svalovej rigidity.
Caudate nucleus a globus pallidus sa podieľajú na takých integračných procesoch, ako je podmienená reflexná aktivita a motorická aktivita. Toto sa zisťuje stimuláciou jadra caudatus, putamen a globus pallidus, deštrukciou a zaznamenávaním elektrickej aktivity.
Priama stimulácia niektorých zón nucleus caudatus spôsobí otočenie hlavy opačným smerom ako stimulovaná hemisféra, zviera sa začne pohybovať po kruhu, t.j. dochádza k takzvanej obehovej reakcii.
U ľudí stimulácia jadra caudatus pri neurochirurgickom zákroku narúša rečový kontakt s pacientom: ak pacient niečo povedal, stíchne a po skončení podráždenia si nepamätá, že bol oslovený. V prípadoch poranenia mozgu s podráždením hlavy nucleus caudatus pacienti pociťujú retro-, antero-, retroanterográdnu amnéziu.
Stimulácia nucleus caudatus môže úplne zabrániť vnímaniu bolestivej, zrakovej, sluchovej a inej stimulácie. Podráždenie ventrálnej oblasti nucleus caudate sa znižuje a dorzálna oblasť zvyšuje slinenie.
Pri poškodení nucleus caudatus sa pozorujú výrazné poruchy vyššej nervovej činnosti, sťažená orientácia v priestore, zhoršenie pamäti, spomalený rast tela. Po obojstrannom poškodení nucleus caudatus podmienené reflexy na dlhú dobu miznú, vývoj nových reflexov sa stáva ťažkým, celkové správanie je charakterizované stagnáciou, zotrvačnosťou a ťažkosťami s prepínaním. Pri postihnutí nucleus caudatus sa okrem porúch vyššej nervovej činnosti zaznamenávajú poruchy hybnosti. Mnohí autori poznamenávajú, že u rôznych zvierat sa pri obojstrannom poškodení striata objavuje nekontrolovateľná túžba posunúť sa vpred a pri jednostrannom poškodení dochádza k manéžovým pohybom.
Škrupina je charakterizovaná účasťou na organizácii stravovacieho správania: vyhľadávanie potravy, orientácia v potrave, zachytávanie potravy a jej trávenie pri poruche funkcie škrupiny dochádza k množstvu trofických porúch kože a vnútorných orgánov; Podráždenie škrupiny vedie k zmenám dýchania a slinenia.
Ako už bolo spomenuté, podráždenie jadra caudate inhibuje podmienený reflex vo všetkých štádiách jeho implementácie. Podráždenie jadra caudatus zároveň zabraňuje zániku podmieneného reflexu, t.j. vývoj inhibície; zviera prestáva vnímať nové prostredie. Vzhľadom na to, že stimulácia jadra caudatus vedie k inhibícii podmieneného reflexu, dalo by sa očakávať, že deštrukcia jadra caudatus spôsobí uľahčenie aktivity podmieneného reflexu. Ukázalo sa však, že deštrukcia jadra caudatus vedie aj k inhibícii podmienenej reflexnej aktivity. Zdá sa, že funkcia jadra caudatus nie je jednoducho inhibičná, ale pozostáva z korelácie a integrácie procesov RAM. Potvrdzuje to aj skutočnosť, že informácie z rôznych zmyslových systémov sa zbiehajú na neurónoch nucleus caudatus, keďže väčšina týchto neurónov je polysenzorická.
Bledá lopta má prevažne veľké Golgiho neuróny typu 1. Spojenie medzi globus pallidus a thalamom, putamenom, caudate nucleus, stredným mozgom, hypotalamom a somatosenzorickým systémom naznačuje jeho účasť na organizácii jednoduchých a zložitých foriem správania.
Stimulácia globus pallidus pomocou implantovaných elektród spôsobuje kontrakciu svalov končatín, aktiváciu alebo inhibíciu gama motorických neurónov miechy.
Stimulácia globus pallidus na rozdiel od stimulácie nucleus caudatus nespôsobuje inhibíciu, ale vyvoláva orientačnú reakciu, pohyby končatín, stravovacie správanie (čuchanie, žuvanie, prehĺtanie a pod.).
Poškodenie globus pallidus spôsobuje u ľudí hypomimiu, maskovitý vzhľad tváre, tras hlavy a končatín (a tento chvenie mizne v pokoji, počas spánku a zintenzívňuje sa pohybmi) a monotónnosť reči. Pri poškodení globus pallidus sa pozoruje myoklonus - rýchle zášklby svalov jednotlivých skupín alebo jednotlivých svalov rúk, chrbta, tváre.
V prvých hodinách po poškodení globus pallidus v akútnom experimente na zvieratách sa motorická aktivita prudko znížila, pohyby boli charakterizované nekoordinovanosťou, bola zaznamenaná prítomnosť neúplnej nekoordinácie, neúplné pohyby a pri sedení bola visiaca poloha. Keď sa zviera začalo pohybovať, dlho sa nemohlo zastaviť. U človeka s dysfunkciou globus pallidus je nástup pohybov obtiažny, pri vstávaní miznú pomocné a reaktívne pohyby, narúšajú sa priateľské pohyby paží pri chôdzi, objavuje sa príznak propulzie: dlhodobá príprava na pohyb, potom rýchly pohyb a zastavenie. Takéto cykly sa u pacientov mnohokrát opakujú.
Plot obsahuje polymorfné neuróny rôznych typov. Vytvára spojenie predovšetkým s mozgovou kôrou.
Hlboká lokalizácia a malá veľkosť plotu predstavujú určité ťažkosti pre jeho fyziologické štúdium. Toto jadro je tvarované ako úzky pásik šedej hmoty, ktorý sa nachádza pod mozgovou kôrou hlboko v bielej hmote.
Stimulácia plota vyvoláva indikatívnu reakciu, otáčanie hlavy v smere podráždenia, žuvanie, prehĺtanie, niekedy aj zvracanie pohybov. Podráždenie z plota inhibuje podmienený reflex na svetlo a má malý vplyv na podmienený reflex na zvuk. Stimulácia plota počas jedenia brzdí proces jedenia jedla.
Je známe, že hrúbka plotu ľavej hemisféry u ľudí je o niečo väčšia ako hrúbka pravej; pri poškodení plotu pravej hemisféry sa pozoruje porucha reči.
Bazálne gangliá mozgu sú teda integračnými centrami pre organizáciu motoriky, emócií a vyššej nervovej aktivity a každá z týchto funkcií môže byť posilnená alebo inhibovaná aktiváciou jednotlivých útvarov bazálnych ganglií.
Amygdala leží v bielej hmote spánkového laloku hemisféry, približne 1,5–2 cm za temporálnym pólom. Amygdala (corpus amygdoloideum), amygdala je subkortikálna štruktúra limbického systému, ktorá sa nachádza hlboko v spánkovom laloku mozgu. Neuróny amygdaly majú rôznu formu, funkciu a neurochemické procesy v nich. Funkcie amygdaly sú spojené s poskytovaním obranného správania, autonómnych, motorických, emocionálnych reakcií a motivácie podmieneného reflexného správania.
Elektrická aktivita mandlí je charakterizovaná osciláciami rôznych amplitúd a frekvencií. Rytmy pozadia môžu korelovať s rytmom dýchania a srdcových kontrakcií.
Amygdala reaguje mnohými svojimi jadrami na zrakové, sluchové, interoceptívne, čuchové a kožné podráždenia a všetky tieto podráždenia spôsobujú zmeny v činnosti ktoréhokoľvek z jadier amygdaly, t.j. Jadrá amygdaly sú multisenzorické. Reakcia jadra na vonkajšie podnety trvá spravidla do 85 ms, t.j. podstatne menej ako reakcia na podobnú stimuláciu neokortexu.
Neuróny majú výraznú spontánnu aktivitu, ktorá môže byť zvýšená alebo inhibovaná senzorickou stimuláciou. Mnohé neuróny sú multimodálne a multisenzorické a spúšťajú sa synchrónne s rytmom theta.
Podráždenie jadier amygdaly vytvára výrazný parasympatický účinok na činnosť kardiovaskulárneho a dýchacieho systému, vedie k zníženiu (zriedkavo k zvýšeniu) krvného tlaku, zníženiu srdcovej frekvencie, narušeniu vedenia vzruchu prevodový systém srdca, výskyt arytmie a extrasystoly. V tomto prípade sa vaskulárny tonus nemusí meniť.
Spomalenie rytmu srdcových kontrakcií pri postihnutí mandlí má dlhú latentnú periódu a má dlhodobý účinok
Podráždenie jadier mandlí spôsobuje útlm dýchania a niekedy reakciu kašľa.
Pri umelej aktivácii amygdaly sa objavujú reakcie smrkania, olizovania, žuvania, prehĺtania, slinenia, zmeny peristaltiky tenkého čreva a účinky nastupujú s dlhou latentnou periódou (až 30-45 s po podráždení). Stimulácia mandlí na pozadí aktívnych kontrakcií žalúdka alebo čriev tieto kontrakcie inhibuje.
Rôzne účinky podráždenia mandlí sú spôsobené ich spojením s hypotalamom, ktorý reguluje fungovanie vnútorných orgánov.
Poškodenie amygdaly u zvierat znižuje primeranú prípravu autonómneho nervového systému na organizáciu a realizáciu behaviorálnych reakcií, čo vedie k hypersexualite, vymiznutiu strachu, pokoja a neschopnosti zúriť a agresie. Zvieratá sa stávajú dôverčivými. Napríklad opice s poškodenou amygdalou sa pokojne priblížia k zmiji, ktorá im predtým spôsobovala hrôzu a útek. Zrejme, ak sú mandle poškodené, niektoré vrodené znaky zmiznú nepodmienené reflexy, uvedomujúc si spomienku na nebezpečenstvo.
Biela hmota hemisféry zahŕňa vnútornú kapsulu a vlákna, ktoré majú rôzne smery. Mali by sa rozlišovať tieto typy vlákien: 1) vlákna prechádzajúce do druhej hemisféry mozgu cez jej komisúry (corpus callosum, anterior commissura, fornix commissura) a smerujúce do kôry a bazálnych ganglií druhej strany ( komisurálne vlákna); 2) systémy vlákien spájajúcich oblasti kôry a subkortikálnych centier v jednej polovici mozgu ( asociatívne); 3) vlákna idúce z mozgovej hemisféry do jej základných častí, do miechy a v opačnom smere od týchto útvarov ( projekčné vlákna).
Ďalším úsekom telencephalonu je corpus callosum, ktoré je tvorené komisurálnymi vláknami spájajúcimi obe hemisféry. Voľný horný povrch corpus callosum, smerujúci k pozdĺžnej trhline veľkého mozgu, je pokrytý tenkou platňou šedej hmoty. Stredná časť corpus callosum je jeho kmeň– vpredu sa ohýba smerom nadol, formuje sa koleno corpus callosum, ktorý sa preriedením mení na zobák, pokračujúc smerom nadol do koncová (hraničná) doska. Zhrubnutá zadná časť corpus callosum končí voľne vo forme hrebeňa. Vlákna corpus callosum tvoria jeho vyžarovanie v každej hemisfére veľkého mozgu. Vlákna rodu corpus callosum spájajú kôru predných lalokov pravej a ľavej hemisféry. Vlákna mozgového kmeňa spájajú šedú hmotu parietálneho a temporálneho laloku. Valček obsahuje vlákna spájajúce kôru okcipitálnych lalokov. Oblasti čelných, parietálnych a okcipitálnych lalokov každej hemisféry sú oddelené od corpus callosum drážkou s rovnakým názvom.
Upozorňujeme, že pod corpus callosum je tenká biela doska - trezor, pozostávajúci z dvoch oblúkových prameňov spojených vo svojej strednej časti priečnou komisurou oblúka (obr.). Telo klenby sa postupne v prednej časti vzďaľuje od corpus callosum, klenie sa dopredu a dole a pokračuje do stĺpa klenby. Spodná časť každý stĺp fornixu sa najskôr približuje k terminálnej doske a potom sa stĺpy fornixu rozchádzajú na bočné strany a sú nasmerované nadol a dozadu a končia v mastoidných telách.
Medzi crura fornixu vzadu a terminálnou doskou vpredu je priečnik predná (biela) komisura, ktorý spolu s corpus callosum spája obe hemisféry veľkého mozgu.
Zozadu pokračuje telo fornixu do plochého stopky fornixu, zrasteného so spodným povrchom corpus callosum. Crus fornixu sa postupne posúva laterálne a dole, oddeľuje sa od corpus callosum, stáva sa ešte hustejším a na jednej strane sa spája s hipokampom a vytvára hipokampálnu fimbriu. Voľná strana fimbrie, privrátená k dutine dolného rohu laterálnej komory, končí háčikom, ktorý spája temporálny lalok telencephalon s diencephalon.
Oblasť ohraničená hore a vpredu corpus callosum, dole jeho zobákom, terminálnou platničkou a prednou komizúrou, vzadu crus fornixu, zaberá na každej strane sagitálne umiestnená tenká platňa - priehľadná priehradka. Medzi doskami priehľadného septa je úzka sagitálna dutina s rovnakým názvom, ktorá obsahuje priehľadnú kvapalinu. Lamina pellucidum je stredná stena predného rohu laterálnej komory.
Pozrime sa na štruktúru vnútorná kapsula(capsula internet) - hrubá, šikmá doska bielej hmoty, ohraničená na bočnej strane lentikulárnym jadrom a na mediálnej strane hlavou nucleus caudate (vpredu) a talamom (vzadu). Vnútornú kapsulu tvoria projekčné vlákna spájajúce mozgovú kôru s ostatnými časťami centrálneho nervového systému. Vlákna vzostupných ciest, ktoré sa rozchádzajú v rôznych smeroch do mozgovej kôry, tvoria žiarivá koruna. Smerom nadol sú vlákna zostupných dráh vnútornej kapsuly vo forme kompaktných zväzkov smerované k stopke stredného mozgu.

Obr.9. Fornix a hipokampus.
1 – corpus callosum, 2 – jadro fornixu, 3 – crus fornixu, 4 – predná komizúra, 5 – stĺpik fornixu, 6 – mastoidné telo, 7 – fimbria hipokampu, 8 – uncus, 9 – zubaté gyrus, 10 – parahipocampal gyrus, 11 – hippocampal peduncle, 12 – hippocampus, 13 – laterálna komora (otvorená), 14 – vtáčia ostroha, 15 – fornix commissura.
Upozorňujeme, že dutiny mozgových hemisfér sú postranné komory(I a II), ktoré sa nachádzajú v hrúbke bielej hmoty pod corpus callosum (obr. 11). Každá komora má štyri časti: predný roh leží v prednom laloku, centrálna časť je v parietálnom laloku, zadný roh- v okcipitálnom spodný roh- v spánkovom laloku. Predný roh oboch komôr je oddelený od susednej dvoma platňami priehľadnej priehradky. Centrálna časť laterálnej komory sa ohýba zhora okolo talamu, tvorí oblúk a prechádza zozadu do zadného rohu, smerom nadol do dolného rohu. Stredná stena dolného rohu je hippocampus(časť starovekej kôry), ktorá zodpovedá hlbokej ryhe rovnakého mena na mediálnom povrchu pologule. Fimbria sa tiahne mediálne pozdĺž hipokampu, ktorý je pokračovaním crus fornixu (obr.). Na strednej stene zadného rohu laterálnej komory mozgu je výčnelok - hippocampus, čo zodpovedá kalkarínovej drážke na mediálnom povrchu hemisféry. Choroidný plexus vyčnieva do centrálnej časti a dolného rohu laterálnej komory, ktorá sa cez interventrikulárny foramen spája s choroidálnym plexom tretej komory.

Obr. 10. Projekcia komôr na povrch veľkého mozgu.
1 – frontálny lalok, 2 – centrálny sulcus, 3 – laterálna komora, 4 – okcipitálny lalok, 5 – zadný roh laterálnej komory, 6 – IV komora, 7 – mozgový akvadukt, 8 – III komora, 9 – centrálna časť komory postranná komora, 10 – dolný roh postrannej komory, 11 – predný roh postrannej komory.

Obr. Predná časť mozgu na úrovni centrálnej časti laterálnych komôr.
1 – centrálna časť laterálnej komory, 2 – plexus cievnatky laterálnej komory, 3 – predná vilózna artéria, 4 – vnútorná mozgová žila, 5 – fornix, 6 – corpus callosum, 7 – cievna báza tretej komory, 8– cievnatka plexus tretej komory, 9 – III komora, 10 – talamus, 11 – pripojená platnička, 12 – talamostriatálna žila, 13 – nucleus caudatus.
Bazálne (subkortikálne) jadrá (nuclei basales) mozgu sa nachádzajú pod bielou hmotou vo vnútri predného mozgu, hlavne v predných lalokoch. Medzi bazálne jadrá patrí caudatus nucleus (nucleus caudatus), putamen (putamen), claustrum a globus pallidus (globus pallidus).
Caudate jadro. Shell
Jadro caudatus (nucleus caudatus) a schránka (putamen) sú evolučne neskoršími útvarmi ako globus pallidus a funkčne naň pôsobia inhibične.
Caudate nucleus a putamen majú podobnú histologickú štruktúru. Ich neuróny patria do Golgiho buniek typu II, t.j. majú krátke dendrity a tenký axón; ich veľkosť je až 20 mikrónov. Tieto neuróny sú 20-krát početnejšie ako Golgiho neuróny typu I, ktoré majú rozsiahlu sieť dendritov a sú veľké asi 50 mikrónov.
Funkcie akejkoľvek mozgovej formácie sú určené predovšetkým ich spojeniami, ktorých majú bazálne gangliá pomerne veľa. Tieto spojenia majú jasné zameranie a funkčný obrys.
Caudate nucleus a putamen prijímajú zostupné spojenia primárne z extrapyramídového kortexu cez subcallosal fasciculus. Iné oblasti mozgovej kôry tiež posielajú veľké množstvo axónov do nucleus caudate a putamen.
Hlavná časť axónov nucleus caudate a putamen smeruje do globus pallidus, odtiaľ do talamu a až z neho do zmyslových polí. V dôsledku toho medzi týmito formáciami existuje začarovaný kruh spojení. Caudatus nucleus a putamen majú funkčné spojenia aj so štruktúrami ležiacimi mimo tohto kruhu: so substantia nigra, červeným jadrom, Lewisovým telom, vestibulálnymi jadrami, mozočkom, γ -bunkami miechy.
Množstvo a povaha spojení medzi caudate nucleus a putamen naznačuje ich účasť na integračných procesoch, organizácii a regulácii pohybov a regulácii práce vegetatívnych orgánov.
Podráždenie poľa 8 mozgovej kôry spôsobuje excitáciu neurónov v nucleus caudate a pole 6 spôsobuje excitáciu neurónov v jadre caudate a putamen. Jediná stimulácia senzomotorickej oblasti mozgovej kôry môže spôsobiť excitáciu alebo inhibíciu aktivity neurónov v nucleus caudatus. Tieto reakcie sa vyskytujú v priebehu 10-20 ms, čo poukazuje na priame a nepriame spojenia medzi mozgovou kôrou a nucleus caudatus.
Mediálne jadrá talamu majú priame spojenie s nucleus caudatus, o čom svedčí reakcia jeho neurónov, ku ktorej dochádza 2-4 ms po stimulácii talamu.
Reakcia neurónov v nucleus caudatus je spôsobená podráždením kože, svetelnými a zvukovými podnetmi.
V interakciách medzi nucleus caudate a globus pallidus prevládajú inhibičné vplyvy. Ak je stimulované jadro caudatus, väčšina neurónov globus pallidus je inhibovaná a menšia časť je excitovaná. Ak dôjde k poškodeniu jadra caudatus, zviera vyvíja motorickú hyperaktivitu.
Interakcia substantia nigra a nucleus caudate je založená na priamych a spätnoväzbových spojeniach medzi nimi. Zistilo sa, že stimulácia caudatus nucleus zvyšuje aktivitu neurónov v substantia nigra. Stimulácia substantia nigra vedie k zvýšeniu a deštrukcia vedie k zníženiu množstva dopamínu v jadre caudatus. Zistilo sa, že dopamín sa syntetizuje v bunkách substantia nigra a potom sa transportuje rýchlosťou 0,8 mm/h do synapsií neurónov v nucleus caudatus. V caudate nucleus v 1 g nervové tkanivo hromadí sa až 10 mcg dopamínu, čo je 6-krát viac ako v iných častiach predného mozgu, globus pallidus, a 19-krát viac ako v mozočku. Vďaka dopamínu sa objavuje dezinhibičný mechanizmus interakcie medzi nucleus caudatus a globus pallidus.
Pri nedostatku dopamínu v nucleus caudatus (napríklad pri dysfunkcii substantia nigra) dochádza k dezinhibícii globus pallidus, aktivácii systémov chrbtice a drieku, čo vedie k motorickým poruchám v podobe svalovej rigidity.
Kortikostriálne spojenia sú lokálne lokalizované. Predné oblasti mozgu sú teda spojené s hlavou nucleus caudate. Patológia vznikajúca v jednej zo vzájomne prepojených oblastí kôry – nucleus caudatus – je funkčne kompenzovaná zachovanou štruktúrou.
Caudate nucleus a globus pallidus sa podieľajú na takých integračných procesoch, ako je podmienená reflexná aktivita a motorická aktivita. Toto sa zisťuje stimuláciou jadra caudatus, putamen a globus pallidus, deštrukciou a zaznamenávaním elektrickej aktivity.
Priame podráždenie niektorých zón caudatus nucleus spôsobí otočenie hlavy opačným smerom ako je podráždená hemisféra, zviera sa začne pohybovať v kruhu, t.j. dochádza k takzvanej obehovej reakcii.
Podráždenie iných oblastí caudate nucleus a putamen spôsobuje zastavenie všetkých typov ľudskej alebo zvieracej činnosti: orientácia, emocionálna, motorická, potravinová. Súčasne sa v mozgovej kôre pozoruje aktivita pomalých vĺn.
U ľudí stimulácia jadra caudatus pri neurochirurgickom zákroku narúša rečový kontakt s pacientom: ak pacient niečo povedal, stíchne a po skončení podráždenia si nepamätá, že bol oslovený. V prípadoch poranenia mozgu s podráždením hlavy nucleus caudatus pacienti pociťujú retro-, antero- alebo retroanterográdnu amnéziu.
U zvierat, ako sú opice, stimulácia caudatus nucleus k rôzne štádiá implementácia podmieneného reflexu vedie k inhibícii vykonávania tohto reflexu. Napríklad, ak je kaudátne jadro opice stimulované cez implantované elektródy pred vydaním podmieneného signálu, potom opica nereaguje na signál, akoby ho nepočula; podráždenie jadra po tom, čo opica ide na kŕmidlo na signál alebo už začína odoberať potravu z kŕmidla, vedie k zastaveniu zvieraťa po ukončení podráždenia sa opica bez dokončenia podmienenej reakcie vráti na svoje miesto, t.j. „zabudne“, že to bol dráždivý signál (retrográdna amnézia).
Stimulácia nucleus caudatus môže úplne zabrániť vnímaniu bolestivej, zrakovej, sluchovej a inej stimulácie. Podráždenie ventrálnej oblasti nucleus caudate sa znižuje a dorzálna oblasť zvyšuje slinenie.
Pri stimulácii caudatus nucleus sa predlžujú latentné periódy reflexov a dochádza k narušeniu alterácie podmienených reflexov. Rozvoj podmienených reflexov na pozadí stimulácie jadra caudatus sa stáva nemožným. Zrejme sa to vysvetľuje tým, že stimulácia nucleus caudatus spôsobuje inhibíciu aktivity mozgovej kôry.
Množstvo subkortikálnych štruktúr tiež dostáva inhibičný vplyv nucleus caudate. Stimulácia jadier caudatus teda spôsobila fusiformnú aktivitu v optickom talame, globus pallidus, subtalamickom tele, substantia nigra atď.
Špecifické pre podráždenie caudatus nucleus je teda predovšetkým inhibícia aktivity mozgovej kôry, subkortikálnych útvarov, inhibícia nepodmieneného a podmieneného reflexného správania.
Zároveň sa pri podráždení caudatus nucleus môžu objaviť niektoré typy izolovaných pohybov. Zdá sa, že caudate nucleus má spolu s inhibičnými a excitačnými štruktúrami.
Vypnutie caudatus nucleus je sprevádzané rozvojom hyperkinézy ako sú mimovoľné reakcie tváre, tremor, atetóza, torzný spazmus, chorea (zášklby končatín a trupu ako pri nekoordinovanom tanci), motorická hyperaktivita v podobe bezcieľneho pohybu z miesto na miesto.
Pri poškodení nucleus caudatus sa pozorujú výrazné poruchy vyššej nervovej činnosti, sťažená orientácia v priestore, zhoršenie pamäti, spomalený rast tela. Po obojstrannom poškodení nucleus caudatus podmienené reflexy na dlhú dobu miznú, vývoj nových reflexov sa stáva ťažkým, celkové správanie je charakterizované stagnáciou, zotrvačnosťou a ťažkosťami s prepínaním. U opíc po jednostrannom poškodení caudatus nucleus sa podmienená reakcia obnovila po 30-50 dňoch, predĺžili sa latentné periódy reflexov a objavili sa intersignálne reakcie. Obojstranné poškodenie viedlo k úplnej inhibícii podmienených reflexov. Obojstranné poškodenie zjavne vyčerpáva symetrické kompenzačné mechanizmy.
Pri postihnutí nucleus caudatus sa okrem porúch vyššej nervovej činnosti zaznamenávajú poruchy hybnosti. Mnohí autori poznamenávajú, že u rôznych zvierat sa pri obojstrannom poškodení striata objavuje nekontrolovateľná túžba posunúť sa vpred a pri jednostrannom poškodení dochádza k manéžovým pohybom.
Napriek veľkej funkčnej podobnosti medzi caudate nucleus a putamen existuje množstvo funkcií špecifických pre putamen.
Evolučne sa škrupina objavuje skôr ako caudate nucleus (jeho základy sú už prítomné v rybách).
Škrupina je charakterizovaná účasťou na organizácii stravovacieho správania: hľadanie potravy, orientácia v potrave, zachytávanie potravy a držba potravy; pri poruche funkcie škrupiny sa vyskytuje množstvo trofických porúch kože a vnútorných orgánov (napríklad hepatolentikulárna degenerácia). Podráždenie škrupiny vedie k zmenám dýchania a slinenia.
Ako už bolo spomenuté, podráždenie jadra caudate inhibuje podmienený reflex vo všetkých štádiách jeho implementácie. Podráždenie kaudatového jadra zároveň bráni zániku podmieneného reflexu, t.j. rozvoju inhibície; zviera prestáva vnímať nové prostredie. Vzhľadom na to, že stimulácia jadra caudatus vedie k inhibícii podmieneného reflexu, dalo by sa očakávať, že deštrukcia jadra caudatus spôsobí uľahčenie aktivity podmieneného reflexu. Ukázalo sa však, že deštrukcia jadra caudatus vedie aj k inhibícii podmienenej reflexnej aktivity. Zdá sa, že funkcia caudatus nucleus nie je jednoducho inhibičná, ale spočíva v korelácii a integrácii procesov RAM. Potvrdzuje to aj skutočnosť, že informácie z rôznych zmyslových systémov sa zbiehajú na neurónoch nucleus caudatus, keďže väčšina týchto neurónov je polysenzorická.
Bledá lopta
Globus pallidus (globus pallidus s. pallidum) má prevažne veľké Golgiho neuróny typu I. Prepojenia medzi globus pallidus a thalamom, putamenom, caudate nucleus, stredným mozgom, hypotalamom, somatosenzorickým systémom atď. naznačujú jeho účasť na organizácii jednoduchých a zložitých foriem správania.
Stimulácia globus pallidus pomocou implantovaných elektród spôsobuje kontrakciu svalov končatín, aktiváciu alebo inhibíciu γ-motoneurónov miechy. U pacientov s hyperkinézou podráždenie rôznych častí globus pallidus (v závislosti od miesta a frekvencie podráždenia) zvýšilo alebo znížilo hyperkinézu.
Stimulácia globus pallidus na rozdiel od stimulácie nucleus caudatus nespôsobuje inhibíciu, ale vyvoláva orientačnú reakciu, pohyby končatín, stravovacie správanie (čuchanie, žuvanie, prehĺtanie a pod.).
Poškodenie globus pallidus spôsobuje u ľudí hypomimiu, maskovitý vzhľad tváre, tras hlavy a končatín (a tento chvenie mizne v pokoji, počas spánku a zintenzívňuje sa pohybmi) a monotónnosť reči. Pri poškodení globus pallidus sa pozoruje myoklonus - rýchle zášklby svalov jednotlivých skupín alebo jednotlivých svalov rúk, chrbta, tváre.
V prvých hodinách po poškodení globus pallidus v akútnom experimente na zvieratách sa motorická aktivita prudko znížila, pohyby boli charakterizované nekoordinovanosťou, bola zaznamenaná prítomnosť neúplných pohybov a pri sedení sa vyskytla ovisnutá poloha. Keď sa zviera začalo pohybovať, dlho sa nemohlo zastaviť. U človeka s dysfunkciou globus pallidus je nástup pohybov obtiažny, pri vstávaní miznú pomocné a reaktívne pohyby, narúšajú sa priateľské pohyby paží pri chôdzi, objavuje sa príznak propulzie: dlhodobá príprava na pohyb, potom rýchly pohyb a zastavenie. Takéto cykly sa u pacientov mnohokrát opakujú.
Klaustrum obsahuje polymorfné neuróny rôznych typov. Vytvára spojenie predovšetkým s mozgovou kôrou.
Hlboká lokalizácia a malá veľkosť plotu predstavujú určité ťažkosti pre jeho fyziologické štúdium. Toto jadro je tvarované ako úzky pásik šedej hmoty, ktorý sa nachádza pod mozgovou kôrou hlboko v bielej hmote.
Stimulácia plota vyvoláva indikatívnu reakciu, otáčanie hlavy v smere podráždenia, žuvanie, prehĺtanie, niekedy aj zvracanie pohybov. Podráždenie z plota inhibuje podmienený reflex na svetlo a má malý vplyv na podmienený reflex na zvuk. Stimulácia plota počas jedenia brzdí proces jedenia jedla.
Je známe, že hrúbka plotu ľavej hemisféry u ľudí je o niečo väčšia ako hrúbka pravej; Keď je poškodený plot pravej hemisféry, pozorujú sa poruchy reči.
Bazálne gangliá mozgu sú teda integračnými centrami pre organizáciu motoriky, emócií a vyššej nervovej aktivity a každá z týchto funkcií môže byť posilnená alebo inhibovaná aktiváciou jednotlivých útvarov bazálnych ganglií.
Subkortikálne jadrá (nucl. subcorticales) sa nachádzajú hlboko v bielej hmote hemisfér. Patria sem kaudátne, lentikulárne, amygdaloidné jadrá a plot (obr. 476). Tieto jadrá sú od seba oddelené vrstvami bielej hmoty, ktoré tvoria vnútorné, vonkajšie a vonkajšie kapsuly. Horizontálny rez mozgom ukazuje striedanie bielej a šedej hmoty subkortikálnych jadier.
Topograficky a funkčne sa kaudátne a lentiformné jadrá spájajú do striata (corpus striatum).
Caudatus nucleus (nucl. caudatus) () má kyjovitý tvar a je zahnutý dozadu. Jeho predná časť je rozšírená, nazývaná hlava (caput) a nachádza sa nad lentikulárnym jadrom, a jeho zadná časť - chvost (cauda) prechádza nad a bočne k talamu, oddelený od neho medulárnymi pruhmi (stria medullaris). Hlava nucleus caudatus sa podieľa na tvorbe laterálnej steny predného rohu laterálnej komory (cornu anterius ventriculi lateralis). Jadro kaudátu pozostáva z malých a veľkých pyramídových buniek. Medzi lentiformným a kaudátovým jadrom je vnútorná kapsula (capsula interna).
Lentiformné jadro (nucl. lentiformis) sa nachádza laterálne a vpredu od talamu. Má klinovitý tvar s vrcholom smerujúcim k stredovej čiare. Medzi zadným okrajom lentikulárneho jadra a talamom je zadná noha vnútorného puzdra (crus posterius capsulae internae) (obr. 476). Predná strana lentiformného jadra zospodu a vpredu je zrastená s hlavou nucleus caudatus. Dva pruhy bielej hmoty oddeľujú nukl. lentiformis na tri segmenty: bočný segment - škrupina (putamen), ktorá má tmavšiu farbu, je umiestnená na vonkajšej strane a dve staré časti bledej gule (globus pallidus) kužeľovitého tvaru sú obrátené do stredu.
476. Horizontálny rez veľkým mozgom.
1 - genu corporis callosi; 2 - hlavička č. caudati; 3 - crus anterius capsulae internae; 4 - vonkajšia kapsula; 5 - klaustrum; 6 - kapsula extréma; 7 - ostrovček; 8 - putamen; 9 - globus pallidus; 10 - crus posterius; 11 - talamus; 12 - plexus chorioideus; 13 - cornu posterius ventriculi lateralis; 14 - sulcus calcarinus; 15 - vermis cerebelli; 16 - splenium corporis callosi; 17 - tr. n. cochlearis et optici; 18 - tr. occipitopontinus et temporopontinus; 19 - tr. thalamocorticalis; 20 - tr. corticospinalis; 21 - tr. corticonuclearis; 22 - tr. frontopontinus.
Klaustrum je tenká vrstva šedej hmoty oddelená vonkajšou kapsulou bielej hmoty od lentikulárneho jadra. Plot nižšie je v kontakte s jadrami prednej perforovanej substancie (substantia perforata anterior).
Jadro amygdaly (corpus amygdaloideum) je skupina jadier a je lokalizovaná vo vnútri predného pólu temporálneho laloku, laterálne od substancie septum perforatum. Toto jadro je možné vidieť iba v prednej časti mozgu.