Faz tecido condutor. Tecidos condutores e educacionais
Os tecidos condutores servem para transportar substâncias dissolvidas na água por toda a planta. nutrientes. Eles surgiram como consequência da adaptação das plantas à vida terrestre. Em conexão com a vida em dois ambientes - solo e ar, surgiram dois tecidos condutores, através dos quais as substâncias se movem em duas direções.
As substâncias sobem ao longo do xilema das raízes às folhas nutrição do solo– água e sais minerais nela dissolvidos (corrente ascendente ou de transpiração).
As substâncias formadas durante a fotossíntese, principalmente a sacarose, movem-se através do floema das folhas às raízes (corrente descendente). Uma vez que estas substâncias são produtos de assimilação dióxido de carbono, o transporte de substâncias através do floema é denominado fluxo de assimilados.
Os tecidos condutores formam um sistema ramificado contínuo no corpo da planta, conectando todos os órgãos - desde as raízes mais finas até os brotos mais jovens. Xilema e floema são tecidos complexos, incluem elementos heterogêneos - condutores, mecânicos, de armazenamento, excretores. Os mais importantes são os elementos condutores, pois desempenham a função de substâncias condutoras.
O xilema e o floema são formados a partir do mesmo meristema e, portanto, estão sempre localizados lado a lado na planta. Os tecidos condutores primários são formados a partir do meristema lateral primário - procâmbio, secundário - a partir do meristema lateral secundário - câmbio. Os tecidos condutores secundários têm uma estrutura mais complexa que os primários.
Xilema (madeira) consiste em elementos condutores - traqueídeos e vasos (traqueias), elementos mecânicos - fibras de madeira (fibras libriformes) e elementos do tecido principal - parênquima lenhoso.
Os elementos condutores do xilema são chamados de elementos traqueais. Existem dois tipos de elementos traqueais – traqueídeos e segmentos vasculares (Fig. 3.26).
A traqueíde é uma célula altamente alongada com paredes primárias intactas. A movimentação das soluções ocorre por filtração através de poros delimitados. Um vaso consiste em muitas células chamadas segmentos de vasos. Os segmentos estão localizados um acima do outro, formando um tubo. Entre segmentos adjacentes do mesmo vaso existem orifícios passantes - perfurações. As soluções movem-se através dos vasos com muito mais facilidade do que através dos traqueídeos.
Arroz. 3.26. Esquema da estrutura e combinação de traqueídeos (1) e segmentos de vasos (2).
Os elementos traqueais em estado maduro e funcional são células mortas que não possuem protoplastos. A preservação dos protoplastos impediria o movimento das soluções.
Vasos e traqueídeos transmitem soluções não apenas verticalmente, mas também horizontalmente para elementos traqueais vizinhos e para células vivas. Paredes laterais traqueídeos e vasos permanecem finos em uma área maior ou menor. Ao mesmo tempo, apresentam espessamentos secundários que conferem resistência às paredes. Dependendo da natureza dos espessamentos das paredes laterais, os elementos traqueais são denominados anulares, espirais, reticulares, escalariformes e porosos pontilhados (Fig. 3.27).

Arroz. 3.27. Tipos de espessamento e porosidade das paredes laterais dos elementos traqueais: 1 – em forma de anel, 2-4 – espiral, 5 – espessamento em tela; 6 – escada, 7 – oposto, 8 – porosidade regular.
Os espessamentos anulares e espirais secundários são fixados à fina parede primária por meio de uma projeção estreita. Quando os espessamentos se unem e se formam pontes entre eles, surge um espessamento reticulado, transformando-se em poros delimitados. Esta série (Fig. 3.27) pode ser considerada uma série morfogenética e evolutiva.
Os espessamentos secundários das paredes celulares dos elementos traqueais tornam-se lignificados (impregnados com lignina), o que lhes confere resistência adicional, mas limita a possibilidade de crescimento em comprimento. Portanto, na ontogênese de um órgão, aparecem primeiro elementos em forma de anel e espirais, ainda capazes de se esticar, que não interferem no crescimento do órgão em comprimento. Quando o crescimento de um órgão cessa, aparecem elementos incapazes de alongamento longitudinal.
No processo de evolução, os traqueídeos apareceram primeiro. Eles são encontrados nas primeiras plantas terrestres primitivas. Os vasos apareceram muito mais tarde, transformando os traqueídeos. Quase todas as angiospermas possuem vasos. As plantas de esporos e gimnospermas, via de regra, não possuem vasos sanguíneos e possuem apenas traqueídeos. Apenas como rara exceção, foram encontrados vasos em esporos como Selaginella, algumas cavalinhas e samambaias, bem como em algumas gimnospermas (Gnetaceae). Porém, nessas plantas, os vasos surgiram independentemente dos vasos das angiospermas. O aparecimento de vasos nas angiospermas marcou uma importante conquista evolutiva, pois facilitou a condução da água; As angiospermas revelaram-se mais adaptadas à vida terrestre.
O parênquima lenhoso e as fibras lenhosas desempenham funções de armazenamento e suporte, respectivamente.
Floema (bast) consiste em elementos condutores - peneira -, células acompanhantes (células companheiras), elementos mecânicos - fibras do floema (bast) e elementos do tecido principal - parênquima do floema (bast).
Ao contrário dos elementos traqueais, os elementos condutores do floema permanecem vivos mesmo no estado maduro, e suas paredes celulares permanecem primárias, não lignificadas. Nas paredes dos elementos crivados existem grupos de pequenos orifícios passantes - campos crivados, através dos quais os protoplastos das células vizinhas se comunicam e ocorre o transporte de substâncias. Existem dois tipos de elementos de peneira - células de peneira e segmentos de tubo de peneira.
As células peneira são mais primitivas; são características de plantas com esporos e gimnospermas. Uma célula de peneira é uma célula única, de comprimento muito alongado, com extremidades pontiagudas. Seus campos de peneira estão espalhados pelas paredes laterais. Além disso, as células da peneira possuem outras características primitivas: carecem de células acompanhantes especializadas e contêm núcleos no estado maduro.
Nas angiospermas, o transporte dos assimilados é realizado por tubos peneirados (Fig. 3.28). Eles consistem em muitas células individuais - segmentos localizados um acima do outro. Os campos crivados de dois segmentos adjacentes formam uma placa crivada. As placas crivadas têm uma estrutura mais perfeita do que os campos crivados (as perfurações são maiores e em maior número).
No estado maduro, os segmentos dos tubos crivosos não possuem núcleo, mas permanecem vivos e conduzem substâncias ativamente. Um papel importante no transporte de assimilados através de tubos de peneira pertence às células acompanhantes (células companheiras). Cada segmento do tubo crivado e sua célula acompanhante (ou duas ou três células no caso de divisão adicional) surgem simultaneamente de uma célula meristemática. As células companheiras possuem núcleos e citoplasma com numerosas mitocôndrias; metabolismo intensivo ocorre neles. Existem numerosas conexões citoplasmáticas entre os tubos da peneira e as células adjacentes a eles. Acredita-se que as células companheiras, juntamente com segmentos de tubos crivados, constituem um único sistema fisiológico que realiza o fluxo de assimilados.

Arroz. 3.28. Floema de caule de abóbora em corte longitudinal (A) e transversal (B): 1 – segmento de tubo peneira; 2 – placa peneira; 3 – célula acompanhante; 4 – parênquima floemático; 5 – placa da peneira entupida.
A duração da operação dos tubos peneirados é curta. Para plantas anuais e brotos acima do solo de gramíneas perenes - não mais do que uma estação de crescimento, para arbustos e árvores - não mais do que três a quatro anos. Quando o conteúdo vivo do tubo crivado morre, a célula companheira também morre.
O parênquima bast consiste em células vivas de paredes finas. Suas células freqüentemente acumulam substâncias de reserva, bem como resinas, taninos, etc. As fibras liberianas desempenham um papel de apoio. Eles não estão presentes em todas as plantas.
No corpo da planta, o xilema e o floema estão localizados lado a lado, formando camadas ou fios separados, chamados de feixes vasculares. Existem vários tipos de feixes condutores (Fig. 3.29).
Os feixes fechados consistem apenas em tecidos vasculares primários, não possuem câmbio e não engrossam ainda mais. Cachos fechados são característicos de plantas com esporos e monocotiledôneas. Os cachos abertos apresentam câmbio e são capazes de espessamento secundário. São característicos das gimnospermas e plantas dicotiledôneas.
Dependendo da posição relativa do floema e do xilema no feixe, os seguintes tipos são diferenciados. Os mais comuns são os feixes colaterais, nos quais o floema fica em um lado do xilema. Os feixes colaterais podem ser abertos (caules de dicotiledôneas e gimnospermas) e fechados (caules de monocotiledôneas). Se com dentro Uma vertente adicional do floema está localizada no xilema; esse feixe é chamado bicolateral; Os feixes bicolaterais só podem ser abertos; são característicos de algumas famílias de plantas dicotiledôneas (abóbora, beladona, etc.).
Existem também feixes concêntricos nos quais um tecido condutor envolve outro. Eles só podem ser fechados. Se houver floema no centro do feixe e o xilema o circundar, o feixe é denominado centrifloema ou anfivasal. Esses feixes são frequentemente encontrados nos caules e rizomas das monocotiledôneas. Se o xilema estiver localizado no centro do feixe e for circundado pelo floema, o feixe é denominado centoxilema ou anficribral. Feixes de centoxilema são comuns em samambaias.

Arroz. 3.29. Tipos de feixes de condução: 1 – colaterais abertos; 2 – bicolateral aberto; 3 – garantias fechadas; 4 – centrifloema concêntrico fechado; 5 – centroxilema concêntrico fechado; K – câmbio; X – xilema; F – floema.
5. Mecânico, armazenamento, tecido com ar. Estrutura, funções
Tecido mecânico- um tipo de tecido de um organismo vegetal, fibras de células vivas e mortas com parede celular altamente espessada, conferindo resistência mecânica ao corpo. Surge do meristema apical, bem como da atividade do procâmbio e do câmbio.
O grau de desenvolvimento dos tecidos mecânicos depende em grande parte das condições; eles estão quase ausentes nas plantas de florestas úmidas e em muitas plantas costeiras, mas estão bem desenvolvidos na maioria das plantas de habitats áridos;
Os tecidos mecânicos estão presentes em todos os órgãos da planta, mas são mais desenvolvidos ao longo da periferia do caule e na parte central da raiz.
Os seguintes tipos de tecidos mecânicos são diferenciados:
colênquima é um tecido elástico de suporte da casca primária de caules jovens de plantas dicotiledôneas, bem como de folhas. Consiste em células vivas com membranas primárias não lignificadas e desigualmente espessadas, alongadas ao longo do eixo do órgão. Fornece suporte para a planta.
O esclerênquima é um tecido durável feito de células que morrem rapidamente com membranas lignificadas e uniformemente espessadas. Fornece força aos órgãos e a todo o corpo das plantas. Existem dois tipos de células do esclerênquima:
as fibras são células longas e finas, geralmente coletadas em fios ou feixes (por exemplo, fibra ou fibras de madeira).
esclereídes são células mortas redondas com membranas lignificadas muito espessas. Eles formam o tegumento, cascas de nozes, sementes de cerejas, ameixas, damascos; conferem à polpa das peras o seu carácter grosseiro característico. Encontrado em grupos na crosta de coníferas e alguns madeira dura, nas cascas duras de sementes e frutos. Suas células Forma redonda com paredes grossas e um núcleo pequeno.
Os tecidos mecânicos fornecem força aos órgãos das plantas. Eles formam uma estrutura que sustenta todos os órgãos da planta, resistindo à fratura, compressão e ruptura. As principais características da estrutura dos tecidos mecânicos, garantindo sua resistência e elasticidade, são o poderoso espessamento e lignificação de suas membranas, o fechamento próximo entre as células e a ausência de perfurações nas paredes celulares.
Os tecidos mecânicos são mais desenvolvidos no caule, onde são representados por fibra e fibras de madeira. Nas raízes, o tecido mecânico está concentrado no centro do órgão.
Dependendo da forma das células, sua estrutura, estado fisiológico e método de espessamento das membranas celulares, distinguem-se dois tipos tecido mecânico: colênquima e esclerênquima (Fig. 8.4).

Arroz. 8.4. Tecidos mecânicos: a - colênquima angular; 6- esclerênquima; c - esclereídes de frutos de ameixa cereja: 1 - citoplasma, 2 - parede celular espessada, 3 - túbulos porosos.
O colênquima é representado por células vivas do parênquima com membranas espessadas de forma irregular, tornando-as especialmente bem adaptadas para fortalecer órgãos jovens em crescimento. Por serem primárias, as células do colênquima se esticam facilmente e praticamente não interferem no alongamento da parte da planta onde estão localizadas. O colênquima geralmente está localizado em filamentos separados ou em um cilindro contínuo sob a epiderme do caule jovem e dos pecíolos das folhas, e também margeia as nervuras das folhas dicotiledôneas. Às vezes, o colênquima contém cloroplastos.
O esclerênquima consiste em células alongadas com membranas uniformemente espessadas, muitas vezes lignificadas, cujo conteúdo morre nos estágios iniciais. As membranas das células do esclerênquima apresentam alta resistência, próxima à resistência do aço. Este tecido está amplamente representado nos órgãos vegetativos das plantas terrestres e forma seu suporte axial.
Existem dois tipos de células do esclerênquima: fibras e esclereídes. As fibras são células longas e finas, geralmente coletadas em fios ou feixes (por exemplo, fibra ou fibras de madeira). As esclereídes são células redondas e mortas com membranas lignificadas muito espessas. Eles formam o tegumento, cascas de nozes, sementes de cerejas, ameixas e damascos; conferem à polpa das peras o seu carácter grosseiro característico.
O tecido fundamental, ou parênquima, consiste em células vivas, geralmente de paredes finas, que formam a base dos órgãos (daí o nome tecido). Contém componentes mecânicos, condutores e outros tecidos permanentes. O tecido principal desempenha uma série de funções e, portanto, distinguem entre assimilação (clorênquima), armazenamento, pneumático (aerênquima) e parênquima aquífero (Fig. 8.5).

Figura 8.5. Tecidos de parênquima: 1-3 - portadores de clorofila (colunares, esponjosos e dobrados, respectivamente); 4-armazenamento (células com grãos de amido); 5 - pneumático ou aerênquima.
Proteínas, carboidratos e outras substâncias são depositadas nas células do parênquima de armazenamento. Está bem desenvolvido nos caules das plantas lenhosas, nas raízes, tubérculos, bulbos, frutos e sementes. Plantas em habitats desérticos (cactos) e pântanos salgados possuem parênquima aquífero em seus caules e folhas, que serve para acumular água (por exemplo, grandes exemplares de cactos do gênero Carnegia contêm até 2 a 3 mil litros de água em seus tecidos) . As plantas aquáticas e pantanosas desenvolvem um tipo especial de tecido básico - parênquima aéreo ou aerênquima. As células do aerênquima formam grandes espaços intercelulares portadores de ar, através dos quais o ar é fornecido às partes da planta cuja conexão com a atmosfera é difícil.
Aerênquima (ou Erenquima) é um tecido que contém ar nas plantas, construído a partir de células interconectadas de modo que grandes vazios cheios de ar (grandes espaços intercelulares) permaneçam entre elas.
Em alguns manuais, o aerênquima é considerado um tipo de parênquima principal.
O aerênquima é construído a partir de células comuns do parênquima ou de células em forma de estrela conectadas entre si por seus esporões. Caracterizado pela presença de espaços intercelulares.
Finalidade: Esse tecido portador de ar é encontrado em plantas aquáticas e pantanosas, e sua finalidade é dupla. Em primeiro lugar, é um contentor de reservas de ar para as necessidades de troca gasosa. Nas plantas totalmente imersas em água, as condições de troca gasosa são muito menos convenientes do que nas plantas terrestres. Enquanto estas últimas estão rodeadas de ar por todos os lados, as plantas aquáticas são, na melhor das hipóteses, encontradas em ambiente reservas muito pequenas; Essas reservas já são absorvidas pelas células superficiais, e não atingem mais a profundidade dos órgãos grossos. Nessas condições, uma planta pode garantir as trocas gasosas normais de duas maneiras: aumentando a superfície de seus órgãos com a correspondente diminuição de sua massividade, ou coletando reservas de ar dentro de seus tecidos. Ambos os métodos são observados na realidade.
Por um lado, em muitas plantas as folhas subaquáticas são extremamente dissecadas, como, por exemplo, no botão de ouro da água (inglês) russo. (Ranunculus aquatilis), Ouvirandrafene s tralis, etc.
Por outro lado, no caso de órgãos maciços, são uma massa esponjosa solta e cheia de ar. Durante o dia, quando, graças ao processo de assimilação, a planta libera oxigênio muitas vezes mais do que o necessário para a respiração, o oxigênio liberado é coletado em reserva nos grandes espaços intercelulares do aerênquima. Em tempo ensolarado, quantidades significativas de oxigênio liberado não cabem nos espaços intercelulares e saem através de várias aberturas aleatórias nos tecidos. Com o início da noite, quando o processo de assimilação cessa, o oxigênio armazenado é gradativamente consumido para a respiração celular e, em troca, o dióxido de carbono é liberado pelas células nas cavidades aéreas do aerênquima, que por sua vez é utilizado durante o dia para as necessidades de assimilação. Assim, dia e noite, os resíduos da planta, graças à presença do aerênquima, não são desperdiçados, mas ficam em reserva para serem utilizados no próximo período de atividade.
Quanto às plantas pantanosas, suas raízes encontram-se em condições respiratórias particularmente desfavoráveis. Sob uma camada de água, em solo saturado de água, vários tipos processos de fermentação e decomposição; oxigênio no máximo camadas superiores o solo já foi totalmente absorvido, criam-se então as condições para a vida anaeróbica, ocorrendo na ausência de oxigênio. As raízes das plantas do pântano não poderiam existir nessas condições se não tivessem suprimento de ar no aerênquima.
Diferença entre plantas pantanosas e plantas não completamente submersas plantas aquáticas dos totalmente imersos é que a renovação dos gases no interior do aerênquima ocorre não só pela atividade vital dos tecidos, mas também com o auxílio da difusão (e difusão térmica); nos órgãos terrestres, o sistema de espaços intercelulares se abre para fora com uma massa de minúsculas aberturas - estômatos, através dos quais, por difusão, a composição do ar nos espaços intercelulares é equalizada com o ar circundante. Porém, com plantas de tamanhos muito grandes, tal forma de renovação do ar no aerênquima das raízes não seria rápida o suficiente. Assim, por exemplo, em árvores de mangue que crescem ao longo da costa marítima com fundo lamacento, alguns ramos das raízes crescem para cima a partir da lama e carregam suas pontas para o ar, acima da superfície da água, cuja superfície é perfurada por numerosos buracos. Essas “raízes que respiram” visam mais rápida atualização ar no aerênquima das raízes alimentadoras ramificadas no lodo livre de oxigênio do fundo do mar.
Diminuir Gravidade Específica
A segunda tarefa do aerênquima é reduzir a gravidade específica da planta. O corpo da planta é mais pesado que a água; o aerênquima desempenha o papel de bexiga natatória da planta; graças à sua presença, mesmo órgãos delgados, pobres em elementos mecânicos, ficam retos na água e não caem desordenadamente no fundo. A manutenção dos órgãos, principalmente as folhas, em posição favorável às funções vitais da planta, conseguida nas plantas terrestres ao alto custo de formação de uma massa de elementos mecânicos, é conseguida aqui nas plantas aquáticas simplesmente pelo transbordamento de ar do aerênquima.
Esta segunda tarefa do aerênquima é especialmente expressa nas folhas flutuantes, onde as demandas da respiração poderiam ser satisfeitas sem a ajuda do aerênquima. Graças à abundância de passagens de ar intercelulares, a folha não só flutua na superfície da água, mas também é capaz de suportar algum peso. As folhas gigantes da Vitória Régia são especialmente famosas por esta propriedade. O aerênquima, que atua como bexiga natatória, muitas vezes forma inchaços semelhantes a bolhas na planta. Essas bolhas são encontradas tanto em plantas com flores (Eichhornia crassipes, Trianea bogotensis) quanto em algas superiores: Sargassum bacciferum. Fucus vesiculosus e outras espécies estão equipadas com bexigas natatórias bem desenvolvidas.
Em biologia, tecido é um grupo de células que possuem estrutura e origem semelhantes, e também desempenham as mesmas funções. Nas plantas, os mais diversos e complexos tecidos se desenvolveram durante o processo de evolução nas angiospermas (plantas com flores). Os órgãos das plantas são geralmente formados por vários tecidos. Existem seis tipos de tecidos vegetais: educacionais, básicos, condutores, mecânicos, tegumentares, secretores. Cada tecido inclui subtipos. Entre os tecidos, assim como dentro deles, existem espaços intercelulares - espaços entre as células.
Tecido educacional
Devido à divisão das células do tecido educativo, a planta aumenta em comprimento e espessura. Nesse caso, algumas células do tecido educacional se diferenciam em células de outros tecidos.
As células do tecido educacional são bastante pequenas, firmemente adjacentes umas às outras, possuem um núcleo grande e uma membrana fina.
O tecido educacional nas plantas é encontrado em cones de crescimento raiz (ponta da raiz) e caule (ápice do caule), ocorre nas bases dos entrenós, e o tecido educacional também compõe câmbio(o que garante o crescimento da espessura do caule).
Células do cone de crescimento da raiz. A foto mostra o processo de divisão celular (divergência cromossômica, dissolução do núcleo).
Parênquima ou tecido moído
O parênquima inclui vários tipos de tecidos. Existem tecidos básicos assimilativos (fotossintéticos), de armazenamento, portadores de água e portadores de ar.
Tecido fotossintético consiste em células contendo clorofila, ou seja, células verdes. Essas células têm paredes finas e contêm um grande número de cloroplastos. Sua principal função é a fotossíntese. O tecido de assimilação constitui a polpa das folhas, faz parte da casca dos caules das árvores jovens e dos caules das gramíneas.

Nas células tecido de armazenamento as reservas de nutrientes se acumulam. Este tecido constitui o endosperma das sementes e faz parte de tubérculos, bulbos, etc. O núcleo do caule, as células internas do caule e da casca da raiz e o pericarpo suculento também geralmente consistem em parênquima de armazenamento.
Parênquima aquífero característica apenas de um número de plantas, geralmente em habitats áridos. A água se acumula nas células desse tecido. O tecido aquífero pode ser encontrado tanto nas folhas (babosa) quanto no caule (cactos).
Tecido de ar característica de plantas aquáticas e pantanosas. Sua peculiaridade é a presença grandes quantidades espaços intercelulares contendo ar. Isso facilita a troca gasosa para a planta quando ela está difícil.
Tecido condutor
A função comum de vários tecidos condutores é conduzir substâncias de um órgão vegetal para outro. Nos troncos das plantas lenhosas, as células do tecido condutor estão localizadas na madeira e no floema. Além disso, na madeira existem vasos (traqueia) e traqueídeos, ao longo do qual a solução aquosa se move das raízes e no floema - tubos de peneira, através do qual as substâncias orgânicas se movem das folhas fotossintéticas.
Vasos e traqueídeos são células mortas. A solução aquosa sobe pelos vasos mais rapidamente do que pelos traqueídeos.
Os tubos de peneira são células vivas, mas anucleadas.
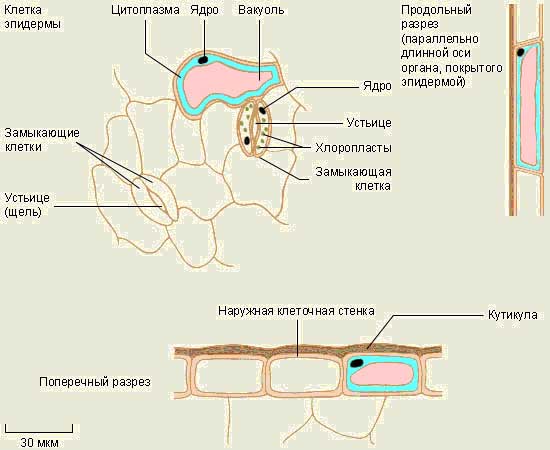
cobrir tecido
O tecido tegumentar inclui a pele (epiderme), cortiça e crosta. A pele cobre as folhas e caules verdes; estas são células vivas; O tampão consiste em células mortas impregnadas com uma substância gordurosa que não permite a passagem de água ou ar.
As principais funções de qualquer tecido tegumentar são proteger as células internas da planta contra danos mecânicos, ressecamento, penetração de microrganismos e mudanças de temperatura.
A cortiça é um tecido de cobertura secundário, pois surge no lugar da casca dos caules e raízes das plantas perenes.
A crosta consiste em cortiça e camadas mortas do tecido principal.
Tecido mecânico
As células dos tecidos mecânicos são caracterizadas por membranas lignificadas altamente espessadas. As funções do tecido mecânico são dar força e elasticidade ao corpo e aos órgãos das plantas.
Nos caules das angiospermas, o tecido mecânico pode estar localizado em uma camada contínua ou em fios separados espaçados uns dos outros.
Nas folhas, as fibras do tecido mecânico geralmente estão localizadas próximas às fibras do tecido condutor. Juntos eles formam as nervuras da folha.
Tecido secretor ou excretor de plantas
As células do tecido secretor secretam substâncias diferentes e, portanto, as funções desse tecido são diferentes. As células excretoras nas plantas revestem as passagens de resina e óleo essencial e formam glândulas e pêlos glandulares peculiares. Os nectários de flores pertencem ao tecido secretor.
As resinas desempenham uma função protetora quando o caule da planta é danificado.
O néctar atrai insetos polinizadores.
Existem células secretoras que removem produtos metabólicos, por exemplo, sais de ácido oxálico.
TECIDOS CONDUTORES
Os tecidos condutores transportam nutrientes em duas direções. Corrente ascendente (transpiração) líquidos ( soluções aquosas e sal) vai junto embarcações E traqueídeos xilema (Fig. 32) desde as raízes até o caule até as folhas e outros órgãos da planta. Corrente descendente (assimilação) a matéria orgânica é transportada das folhas ao longo do caule até os órgãos subterrâneos da planta através

especial tubos de peneira floema (Fig. 33). O tecido condutor de uma planta lembra um pouco sistema circulatório humano, por possuir uma rede axial e radial altamente ramificada; os nutrientes entram em todas as células de uma planta viva. Em cada órgão da planta, o xilema e o floema estão localizados lado a lado e se apresentam na forma de fios - feixes condutores.
Existem tecidos condutores primários e secundários. Os primários diferenciam-se do procâmbio e são formados em órgãos de plantas jovens; os tecidos condutores secundários são mais poderosos e são formados a partir do câmbio.
Xilema (madeira) apresentado traqueídeos E traquéia, ou embarcações.
Traqueídeos- células fechadas alongadas com extremidades recortadas cortadas obliquamente, no estado maduro são representadas por células prosenquimais mortas. O comprimento das células é em média de 1 a 4 mm. A comunicação com os traqueídeos vizinhos ocorre através de poros simples ou delimitados. As paredes são espessadas de forma irregular de acordo com a natureza do espessamento das paredes, os traqueídeos são diferenciados em anulares, espirais, escalariformes, reticulados e porosos (Fig. 34). Os traqueídeos porosos sempre apresentam poros delimitados (Fig. 35). Os esporófitos de todas as plantas superiores possuem traqueídeos e, na maioria das cavalinhas, licófitas, pteridófitas e gimnospermas, eles servem como os únicos elementos condutores do xilema. Traqueídeos


desempenham duas funções principais: condução de água e fortalecimento mecânico do órgão.
Traquéia, ou embarcações, são os principais elementos condutores de água do xilema das angiospermas. As traqueias são tubos ocos constituídos por segmentos individuais; existem buracos nas divisórias entre os segmentos - perfuração, graças ao qual o fluido flui. As traqueias, assim como os traqueídeos, são Sistema fechado: As extremidades de cada traqueia possuem paredes transversais chanfradas com poros delimitados. Os segmentos traqueais são maiores que os traqueídeos: em diâmetro têm cerca de tipos diferentes plantas de 0,1 - 0,15 a 0,3 - 0,7 mm. O comprimento da traquéia varia de vários metros a várias dezenas de metros (para cipós). A traquéia consiste em células mortas, embora nos estágios iniciais de formação estejam vivas. Acredita-se que as traqueias surgiram dos traqueídeos no processo de evolução.
Além da concha primária, a maioria dos vasos e traqueídeos apresentam espessamentos secundários na forma de anéis, espirais, escadas, etc. Espessamentos secundários se formam na parede interna dos vasos (ver Fig. 34). Assim, em um vaso anular, os espessamentos internos das paredes têm a forma de anéis distantes uns dos outros. Os anéis estão localizados transversalmente ao vaso e ligeiramente oblíquos. Em um vaso espiral, a membrana secundária é colocada em camadas do interior da célula na forma de uma espiral; em um vaso reticulado, as áreas não espessadas da concha parecem fendas, lembrando células reticuladas; no vaso escaleno, locais espessados se alternam com outros não espessados, formando uma espécie de escada.
Traqueídeos e vasos - elementos traqueais - estão distribuídos no xilema de diferentes maneiras: em seção transversal em anéis contínuos, formando madeira anelar-vascular, ou espalhados mais ou menos uniformemente por todo o xilema, formando madeira vascular dispersa. A casca secundária é geralmente impregnada com lignina, dando resistência adicional à planta, mas ao mesmo tempo limitando seu crescimento em comprimento.
Além de vasos e traqueídeos, o xilema inclui elementos de viga, consistindo de células formando os raios medulares. Os raios medulares consistem em células vivas do parênquima de paredes finas, através das quais os nutrientes fluem horizontalmente. O xilema também contém células vivas do parênquima lenhoso, que funcionam como transporte de curto alcance e servem como local de armazenamento de substâncias de reserva. Todos os elementos do xilema vêm do câmbio.
Floema- tecido condutor através do qual são transportadas glicose e outras substâncias orgânicas - produtos da fotossíntese das folhas até os locais de sua utilização e deposição (para cones de crescimento, tubérculos, bulbos, rizomas, raízes, frutos, sementes, etc.). O floema também é primário e secundário.

O floema primário é formado a partir do procâmbio, o secundário (floema) - a partir do câmbio. O floema primário carece de raios medulares e de um sistema de elementos crivados menos poderoso que os traqueídeos. Durante a formação do tubo crivado, corpos mucosos aparecem no protoplasto das células - segmentos do tubo crivado, que participam da formação do cordão mucoso próximo às placas crivadas (Fig. 36). Isto completa a formação do segmento do tubo crivado. Os tubos de peneira funcionam na maioria plantas herbáceas uma estação de crescimento e até 3-4 anos para árvores e arbustos. Os tubos de peneira consistem em um número de células alongadas que se comunicam entre si através de septos perfurados - filtro. As cascas dos tubos da peneira em funcionamento não ficam lignificadas e permanecem vivas. As células velhas ficam obstruídas com o chamado corpo caloso e depois morrem e são achatadas sob a pressão das células funcionais mais jovens sobre elas.
Refere-se ao floema parênquima bast, consistindo em células de paredes finas nas quais são depositados nutrientes de reserva. Por raios medulares O floema secundário também realiza transporte de curto alcance de nutrientes orgânicos - produtos da fotossíntese.
Feixes condutores- fios formados, via de regra, por xilema e floema. Se os feixes vasculares são adjacentes aos fios

tecido mecânico (geralmente esclerênquima), então esses feixes são chamados vascular-fibroso. Outros tecidos também podem ser incluídos nos feixes vasculares - parênquima vivo, laticíferos, etc. Os feixes vasculares podem ser completos, quando o xilema e o floema estão presentes, e incompletos, consistindo apenas de xilema (xilema, ou feixe vascular lenhoso) ou floema (floema, ou bast, feixe condutor).
Os feixes vasculares foram originalmente formados a partir do procâmbio. Existem vários tipos de feixes condutores (Fig. 37). Parte do procâmbio pode ser preservada e depois se transformar em câmbio, então o feixe é capaz de espessamento secundário. Esse abrir feixes (Fig. 38). Esses feixes vasculares predominam na maioria das plantas dicotiledôneas e gimnospermas. As plantas com cachos abertos são capazes de crescer em espessura devido à atividade do câmbio, e as áreas lenhosas (Fig. 39, 5) são aproximadamente três vezes maiores que as áreas liberianas (Fig. 39, 5). 2) . Se, durante a diferenciação do feixe vascular do cordão procambial, todo o tecido educativo for totalmente gasto na formação de tecidos permanentes, então o feixe é denominado fechado(Fig. 40). Fechado



feixes vasculares são encontrados nos caules das monocotiledôneas. Madeira e bastão em feixes podem ter posições relativas diferentes. A este respeito, distinguem-se vários tipos de feixes vasculares: colaterais, bicolaterais (Fig. 41), concêntricos e radiais. Garantia, ou lado a lado, - feixes nos quais o xilema e o floema são adjacentes. Bicolateral, ou dupla face, - feixes nos quais dois filamentos do floema se unem lado a lado ao xilema. EM concêntrico em feixes, o tecido do xilema envolve completamente o tecido do floema ou vice-versa (Fig. 42). No primeiro caso, tal pacote é denominado centrifloema. Os feixes de centrofloema estão presentes nos caules e rizomas de algumas plantas dicotiledôneas e monocotiledôneas (begônia, azeda, íris, muitos juncos e lírios). As samambaias os têm. Há também



feixes vasculares intermediários entre os colaterais fechados e os centrifloêmicos. Encontrado nas raízes radial feixes em que a parte central e os raios ao longo dos raios são deixados pela madeira, e cada raio da madeira consiste em vasos centrais maiores, diminuindo gradativamente ao longo dos raios (Fig. 43). Número de raios plantas diferentes não é o mesmo. Entre os raios da madeira existem áreas bastões. Os tipos de feixes condutores são mostrados esquematicamente na Fig. 37. Os feixes vasculares estendem-se ao longo de toda a planta em forma de cordões, que começam nas raízes e percorrem toda a planta ao longo do caule até às folhas e outros órgãos. Nas folhas são chamadas de veias. Sua principal função é conduzir correntes descendentes e ascendentes de água e nutrientes.
O tecido condutor consiste em células alongadas vivas ou mortas que se parecem com tubos.
Os caules e folhas das plantas contêm feixes de tecido condutor. O tecido condutor contém vasos e tubos de peneira.
Embarcações- células ocas mortas conectadas sequencialmente, as partições transversais entre elas desaparecem. Através dos vasos, a água e os minerais dissolvidos nele desde as raízes entram no caule e nas folhas.
Tubos de peneira - células vivas alongadas e sem núcleo, conectadas em série entre si. Através deles, as substâncias orgânicas das folhas (onde foram formadas) se deslocam para outros órgãos da planta.
Tecido condutor garante o transporte de água com minerais dissolvidos nele.
Este tecido forma dois sistemas de transporte:
- para cima(das raízes às folhas);
- para baixo(das folhas a todas as outras partes das plantas).
Ascendente sistema de transporte consiste em traqueídeos e vasos (xilema ou madeira), e os vasos são agentes condutores mais avançados que os traqueídeos.
Nos sistemas descendentes, o fluxo de água com produtos da fotossíntese passa por tubos crivados (floema ou floema).
O xilema e o floema formam feixes vasculares fibrosos - o “sistema circulatório” da planta, que o penetra completamente, conectando-o em um todo.
Os cientistas acreditam que o surgimento dos tecidos está associado na história da Terra ao surgimento das plantas na terra. Quando parte da planta se encontrava no ar e a outra parte (a raiz) no solo, tornou-se necessário levar água e sais minerais das raízes para as folhas, e substâncias orgânicas das folhas para as raízes. Então, no curso da evolução flora Surgiram dois tipos de tecidos condutores - madeira e fibra.
Através da madeira (através de traqueídeos e vasos), a água com minerais dissolvidos sobe das raízes às folhas - esta é uma corrente condutora de água ou ascendente. Através do floema (através de tubos de peneira) as substâncias orgânicas formadas nas folhas verdes fluem para as raízes e outros órgãos da planta - esta é uma corrente descendente.
Tecido educacional
O tecido educacional é encontrado em todas as partes em crescimento da planta. O tecido educacional consiste em células que são capazes de se dividir ao longo da vida da planta. As células aqui ficam muito rapidamente umas sobre as outras. Através da divisão, eles formam muitas células novas, garantindo assim que a planta cresça em comprimento e espessura. As células que aparecem durante a divisão dos tecidos educacionais são então transformadas em células de outros tecidos vegetais.

É o tecido primário a partir do qual todos os outros tecidos vegetais são formados. Consiste em células especiais capazes de múltiplas divisões. São essas células que constituem o embrião de qualquer planta.
Este tecido fica retido na planta adulta. Está localizado:
- na parte inferior do sistema radicular e no topo dos caules (garante o crescimento da planta em altura e o desenvolvimento do sistema radicular) - tecido educacional apical;
- dentro do caule (garante que a planta cresça em largura e engrosse) - tecido educacional lateral.
Ao contrário de outros tecidos, o citoplasma do tecido educacional é mais espesso e denso. A célula possui organelas bem desenvolvidas que proporcionam a síntese de proteínas. O núcleo é caracterizado por um grande tamanho. A massa do núcleo e do citoplasma é mantida em uma proporção constante. O aumento do núcleo sinaliza o início do processo de divisão celular, que ocorre por mitose para as partes vegetativas das plantas e meiose para os meristemas esporogênicos.
No processo de evolução, com o surgimento de plantas superiores em terra, desenvolveram tecidos que atingiram sua maior especialização em plantas com flores. Neste artigo veremos mais de perto o que são os tecidos vegetais, que tipos existem, que funções desempenham, bem como as características estruturais dos tecidos vegetais.
Tecido são grupos de células com estrutura semelhante e que desempenham as mesmas funções.
Os principais tecidos vegetais são mostrados na figura abaixo:
Tipos, funções e estrutura dos tecidos vegetais.


Tecido tegumentar de plantas.


Tecido tegumentar vegetal - crosta
Tecido vegetal condutor.
| Nome do tecido | Estrutura | Localização | Funções |
| 1. Vasos de madeira - xilema | Tubos ocos com paredes lignificadas e conteúdo morto | Madeira (xilema) correndo ao longo da raiz, caule e nervuras das folhas | Conduzindo água e minerais do solo para a raiz, caule, folhas, flores |
|
2. Tubos de peneira de fibra - floema Células acompanhantes ou células companheiras |
Fileira vertical de células vivas com divisórias transversais em forma de peneira Células irmãs de elementos crivados que mantiveram sua estrutura |
Bast (floema), localizado ao longo da raiz, caule e nervuras das folhas Sempre localizado ao longo dos elementos peneirados (acompanha-os) |
Transportando matéria orgânica das folhas ao caule, raiz, flores Participe ativamente no transporte de substâncias orgânicas através dos tubos crivados do floema |
| 3. Condução de feixes fibrosos vasculares | Um complexo de madeira e bastão na forma de fios separados nas gramíneas e uma massa contínua nas árvores | Cilindro central da raiz e caule; veias de folhas e flores | Transportar água e minerais através da madeira; em bast - substâncias orgânicas; fortalecendo órgãos, conectando-os em um único todo |



Tecido mecânico de plantas.

